
The adult digestive system of lacewings changes in a variety of ways during metamorphosis. Additionally, predatory and glyco-pollen feeding lacewings have different characteristics. This post will cover:
- Adult Digestive System, which will talk about the differences between predatory and non-predatory digestive systems, and symbiotic yeast
- Adult Feeding Behaviors, covering migration and flight habits, and responses to infochemicals
Chrysopid Adult Digestion Systems
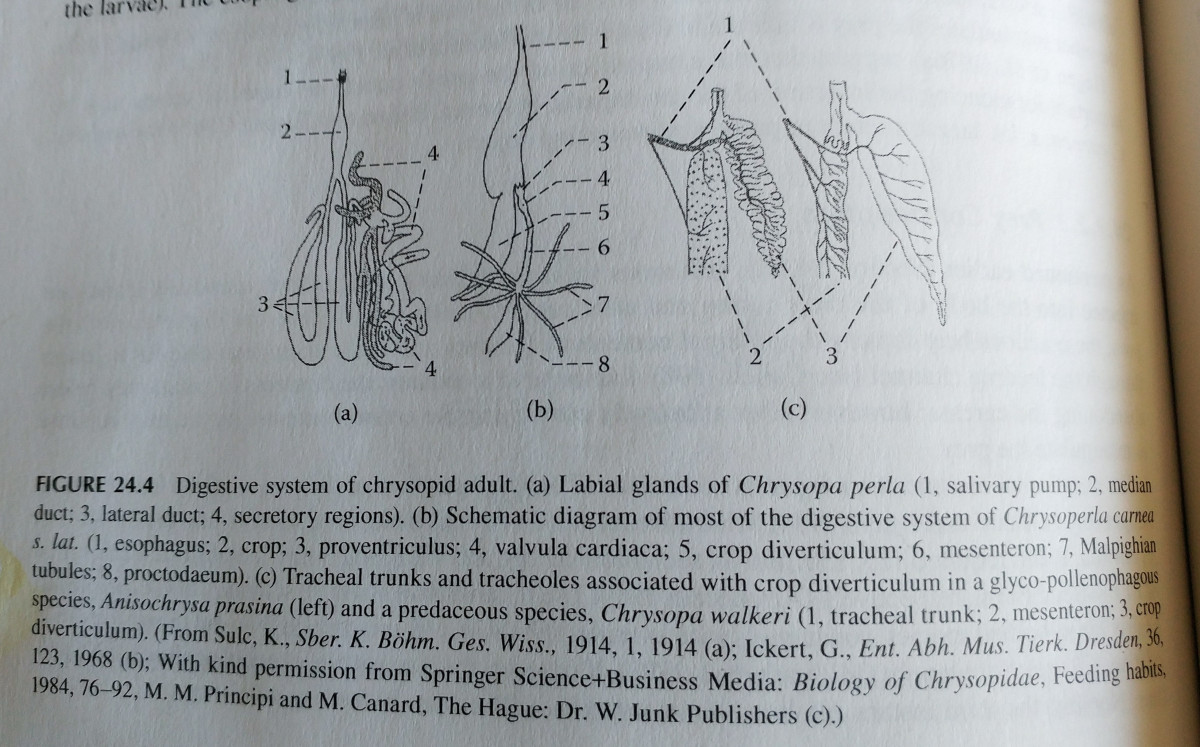 During metamorphosis, adults gain a mouth. The stomodaeum differentiates to a crop and proventriculus, and the proctodaeum opens. Most adults share a common anatomical plan, but there are differences associated with predatory or glyco-pollen diets.
During metamorphosis, adults gain a mouth. The stomodaeum differentiates to a crop and proventriculus, and the proctodaeum opens. Most adults share a common anatomical plan, but there are differences associated with predatory or glyco-pollen diets.
Nonpredaceous Adults
Pollen and nectar feeders have a number of distinct features. They have small, symmetrical mandibles without incisors, spoon shaped lacinia (for holding pollen), and large tracheal trunks that extend to the crop.
Some of these features lack support of association with glyco-pollen feeders. Many structures (the mouthparts especially) lack strong evidence. Some species have large, asymmetrical mandibles , with the left possessing an incisor, which is usually associated with prey feeders.
Many species of glyco-pollen feeders also have symbiotic yeast inside their gut. The symbionts typically live inside the crop diverticulum, which is enlarged and highly folded in adult lacewings. The crop is heavily tracheated, and the trachael trunks extending to it are greatly enlarged. This serves to oxygenate the symbiotic yeast.
Predatory adults lack these features, although some glyco-pollen feeders are exceptions. For example, Hypochrysa elegans (subfamily Nothochrysinae) feed only on pollen, and have large jaws, small tracheal trunks, and no yeast symbionts.
Predaceous Adults
Most adult chrysopids aren’t predatory, and only a small minority of them feed on prey. Out of 75 recognized taxa, only 3 are predaceous. These are Anomalochrysa spp, Atlantochrysa spp, and Chrysopa spp. The first two are restricted to islands and have not been studied extensively. Chrysopa spp, on the other hand, is widely distributed in the Holarctic, and is often used as a beneficial insect in agricultural systems. This species is generally considered carnivorous, however it can feed on pollen, yeast, fungal spores, and honeydew.
There are differentiating features that are assumed to be adapted for this predatory lifestyle. The mandibles are larger and asymmetrical, and have a large tooth or incisor on the left mandible, as well as a molar chewing surface. The lacinia (tip of the maxilla) are unmodified, as they don’t need to catch pollen. The diverticula are still large, however the trachael trunks are smaller, and likely provide less oxygen than those mentioned in glyco-pollen feeders. Additionally, no symbiotic yeast has been discovered in them. It is inconclusive how many of these traits relate to their feeding behaviors, as some glyco-pollen feeders also share similar features.
Symbiotic Yeast
The yeasts found in C. carnea s. lat have been identified as Candida spp, and Metschrikowia spp, and many of these have been found in C. rufilabris and C. comanche. These species are likely quite general among glyco-pollen feeding Chrysopid adults.
The reason for this symbiosis is not yet fully understood. Morphological and experimental evidence suggests that some essential nutrients may be absent or in low concentrations in carbohydrate rich diets. How the yeast is transferred from the crop to the midgut is not concrete, although the most likely scenario is that yeast cells proliferate in the diverticulum, and then the yeast cells or the nutrients they produce are transferred to the midgut, digested, then absorbed. The proventriculus apparently is involved in controlling this transfer.
Gibson & Hunter (2005) questioned the nutritional role of these yeasts, as they couldn’t replicate the results of Hagen et Al (1970)’s resul;ts. This could be because of variation within the C. carnea species complex, or because they were looking for a different species of yeast than those present. Many of these questions have yet to be resolved.
Additionally, how these yeasts are carried throughout generations isn’t completely understood. It could happen after adult emergence, via ingested food. Newer discoveries suggest that vertical transmission, from mother to offspring, may occur. Active yeasts have been found on Chrysopid eggs and within larvae, but it is not clear how they could get these cells from the egg. Larvae pierce the egg with a schlerotized egg burster, and the larval mouth is closed during this stage. Basically, there would be no way for them to ingest it! Perhaps this is part of the reason larvae will cannibalize eggs from their own species?
Adult Feeding Behavior
Most knowledge about feeding behavior is based on a few species, most of which are gylco-pollen feeders. Additionally, habitat and food search is related to the search for oviposition sites.
Postemergence Movement
Adult lacewings are mostly nocturnal, and are generally inactive during the day, hidden beneath leaves. C. carnea s. lat begins its activity at dusk, peaks during early darkness, and becomes inactive at dawn. They usually only fly during the day if they are disturbed, and these flights tend to be short. There are some species that are active during the day, however it is less common.
Lacewing flight generally appears slow, however some species are capable of quick flight and maneuvering. Additionally, lacewings can be carried by wind, especially when they’re more than 5 meters off the ground.
C. carnea displays three types of flight: migratory, appetitive downwind, and appetitive upwind. Migratory flights are long, and lacewings can be dispersed up to 300 km by wind. These flights are closely related to the period of sexual immaturity after emerging from the pupae stage, and generally occur regardless of food nearby. Appetitive downwind begins about 3 days after emergence. Adults begin to take short flights, looking for volatile chemicals from food sources. They typically fly downwind 1-5 m above the vegetation and land when they discover the appropriate odor. If they don’t immediately find the food, they begin appetitive upwind flight, where they move upwind and approach the odor slowly in a series of short flights.
Even after mating (3-4 days post-emergence) and oviposition (5th night), C. carnea keeps moving. The whole population moves downwind every night, which could be to make sure that offspring are well dispersed. Little is known about other species flight patterns.
Infochemicals (volatiles) associated with food (such as prey or honeydew) influence the way lacewings fly. Volatile secondary metabolites may directly act on the predator (these chemicals would acting as synomones), or indirectly, in the feces or honeydew from the prey (kairomones). Additionally, pheromones produced by the prey (acting as kairomones) and volatiles emanating from plants in response to prey damage (synomones) are also attractive.
Attraction to Plant Volatiles
There are many compounds that have been found to be attractive to lacewings.
- Neomatatabiol and other monoterpene alcohols (turpenes) are attractive to Chrysopa formosa and Ch. pallens
- Terpene acetate for C. carnea s lat and Ch. nigricornis
- Caryophyllene, eugenol, 2-phenlethanol, (z)-3-hexenil-acetate
These relationships are beneficial to both the reciever (lacewings, in this case) and the plant, due to reduced pest pressure. The above chemicals are regarded as synomones for habitat location, but if they are assimilated by plant feeders and then excreted as feces, sex pheromones, or honeydew, they are classified as kairomones. This has only been confirmed with neomatatabiol, a synomone made by Actinidia polygama (a plant native to Asia). It has been identified as part of aphid sex pheromones that lacewings react favorably to. The main ingredient is dihydronepetalactol.
Response to Prey Associated Volatiles
Long Distance Attraction
Many lacewings respond to aphid sex pheromones, specifically nepetalactol and dihydronepetalactol, however this is not because of kairomones, but a coincidence in the pheromone chemistry of lacewings and aphids.
Adults may also be attracted to volatiles plants reduce in response to being eaten. So far the only compound that has been verified is methyl salicylate, a response produced by many phloem feeding homopterans. This occurs in many plant species, including some cultivated crops.
Short Distance Response
Volatiles from honeydew cause chrysopids to halt and intensify their search behavior, as well as feeding and oviposition. It’s not known which compounds cause this reaction, and volatiles found in feces is largely unstudied. C. carnea has been observed reacting to dipropyl-disulfide, dimethyldisulfide, allyl-isothiocyanate, and dimethyl-trisulfide. This could be for habitat location (long distance), food (short distance), or a combination of both.
Conclusion
Habitat and food location are an integrated process for lacewings. Food is located by anemochemotactile attraction induced by synomones from plants or kairomones from prey. C. carnea s. lat appears to need a synomone from the plant to react to the kairomone from the prey, so there are many synergistic interactions between compounds. Lacewings likely use a series of compounds from different categories as well as non-chemical, prey related stimuli to locate habitat, food, oviposition sites, and mates.