

Order: Suliformes
Family: Phalacrocoracidae
Genus: Phalacrocorax
Species: Phalacrocorax penicillatus

Breeding plumage in adult
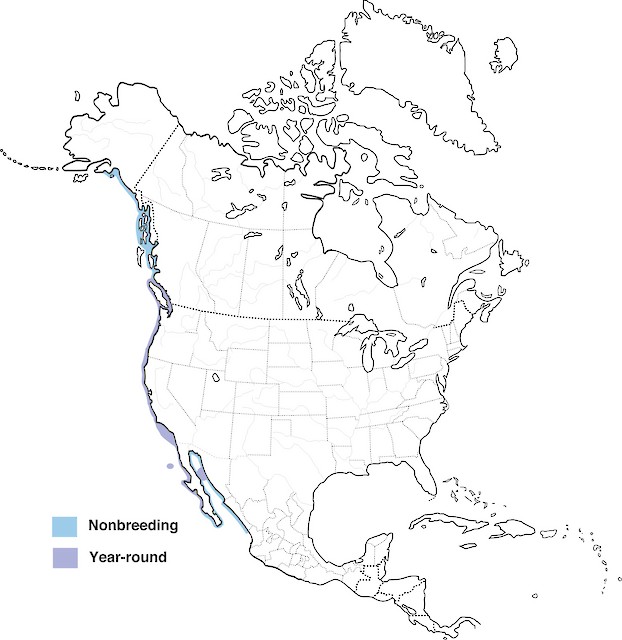
Map by Elizabeth A. Wallace and George E. Wallace
Brandt’s Cormorants are mainly permanent residents in North America, with some local movements. The Cormorants witnessed nesting on Farallon Islands off of California are typically absent in the winter, with high probabilities of going to the adjacent mainland. They only reside in southeastern Alaska during the summer in very small numbers. They sometimes wander along the Mexican coast south of their normal breeding range. These birds are almost never found inland (Audubon, 2017).
These Cormorants are mainly found along inshore coastal waters, especially in areas with kelp beds. They also frequent large bays and occasionally estuaries or coastal lagoons. Their breeding colonies are usually located on gentle slopes on the windward sides of islands, and on the ledges of steep cliffs (North American Bird Conservation Initiative, 2014).
Brandt’s Cormorants are exclusively marine to salt and/or brackish water. They favor rocky shorelines and the open ocean along the Pacific Coast of North America (Wallace & Wallace, 1998). They are common year-round residents in marine subtidal and pelagic zones of California, especially near rocky shores (Garrett and Dunn 1981). These birds even follow salmon runs well upstream on northwestern California rivers, such as Klamath and Smith (McCaskie et al., 1979).
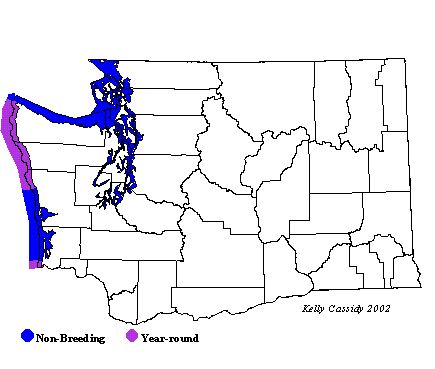
Washington Range Map. Photo by Kelly Cassidy
Brandt’s Cormorants tend to feed on prey on or just above the bottom of bodies of water in rocky areas. They also feed in areas of flat sand or mud (Ainley, Anderson, & Kelly, 1981). Their predominant prey consists mainly of rockfish, anchovies, and damselfish. However they have been observed eating squid if the squid happen to be spawning in their habitat (Ainley et al. 1981). Brandt’s Cormorants have a fairly wide range of dietary preferences that overlap those of both the Pelagic and Double-crested Cormorants. This may be due to Brandt’s having a somewhat large distribution range.
In the Columbia River estuary, Brandt’s Cormorants diets were considerably different from similar studies conducted in other areas. Their prey types were river lamprey, three-spine stickleback, starry flounder, members of the Salmonidae family and Pacific staghorn sculpin. These Brandt’s appeared to be opportunistic feeders. Several species of fish use the Columbia River estuary as a nursery ground, making them seasonally abundant (Couch & Lance, 2004).
They make low, hoarse and guttural croaks, growls, and gargles.
Brandt’s Cormorants are well known divers, reportedly as deep as 240 feet (Palmer 1962). They plunge below the surface and chase prey under water. They grab fish in their bills without spearing it (Wallace & Wallace, 1998).
They breed mostly from March to August, and lay eggs from April to July. They are monogamous, colonial nesters. Clutch sizes typically range from 3-6 and are single-brooded. Incubation is performed by both parents (Marshall & Giles, 1953).
Their nests look like circular drums of dry matter, including items stolen from other nests, grass, moss, weeds, seaweed collected by diving and occasionally sticks or rubbish. Their eggs and are typically pale blue or a blueish white in color. Their hatchlings are born altricial, which means they are considered naked and helpless, requiring extensive parental care (Wallace & Wallace, 1998).

Brandt’s Cormorant nesting at the end of the Breakwater.
Brandt’s Cormorants breed in colonies, with males choosing nest sites and putting on displays to both ward off rivals and attract mates. Their displays include drawing the head back and exposing their blue throat pouch, pointing their bill upward, spreading their tail, and wing fluttering. They also thrust their head forward and downward in rapid repeated gestures (Audubon, 2017).
Bibliography
Ainley, D. G., Anderson, D. W., & Kelly, P. R. (1981). Feeding Ecology of Marine Cormorants in Southwestern North America. The Condor: Ornithological Applications, 83 (2), 120–131.
Audubon. (2017). Brandt’s Cormorant. Audubon Guide to North American Birds.
Carter, H. R., & Hobson, K. A. (1988). Creching Behavior of Brandt’s Cormorant Chicks. The Condor: Ornithological Applications, 90 (2), 395–400.
Cassidy, K. (n.d.). Washington Range Map. 2002. Seattle Audubon Society.
Couch, S. L., & Lance, M. (2004). Diet Composition and Breeding Success of Brandt’s Cormorants in the Columbia River Estuary. Northwestern Naturalist, 85(2), 62-66.
Garrett, K. a. (1981). Birds of southern California. Los Angeles Audubon Society, 480pp.
Kushlan, J. A. (2002). Waterbird Conservation for the Americas: The North American Waterbird Conservation Plan. Washington DC.
Lang Elliott, B. M. (2019). Colony.
Marshall, D. B., & Giles, L. W. (1953). Recent Observations of Birds of Anaho Island. Pyramid Lake, NV: Condor.
McCaskie, G., De Benedictis, P., Erickson, R., & Morlan, J. (1979). Birds of Northern California, An Annotated Field List. Berkeley: Golden Gate Audubon Society.
North American Bird Conservation Initiative. (2014). The State of the Birds 2014 Report. Washington, DC: US Department of Interior.
Nur, N., & Sydeman, W. J. (2009). Survival, Breeding Probability and Reproductive Success in Relation to Population Dynamics of Brandt’s Cormorants Phalacrocorax penicillatus. Bird Study, S92-S103.
Palmer, R. S. (1962). Handbook of North American Birds. New Haven: Yale University Press.
Wallace, E. A., & Wallace, G. E. (1998). Brandt’s Cormorant (Phalacrocorax penicillatus) In The Birds of North America. Cornell Lab of Ornithology, DOI 10.2173.
The populations of Brandt’s Cormorants are stable to declining. Estimates put 151,200 breeding birds on the continent, with a rate of 15 out of 20 on the Continental Concern Score. They are currently listed as a Species of High Concern. However, they have experienced increases in populations from 1900 to 1970 due to legal legislation protecting them from egging and hunting (Kushlan et al. 2002).
To give readers an idea of statistical breeding results, the following example is from a recent study from a bird observatory in Stinson, California:
Males:
- 77% survive to next year (SURGE estimate), of which:
- 28% not observed
- 14% observed, but not breeding
- 35% observed breeding
Females:
- 71% survive to next year (SURGE estimate), of which:
- 34% not observed
- 1% observed, but not breeding
- 36% observed breeding
The factors leading to these results are the adult breeding probabilities, reproductive successes and largely the availability of prey. It’s also speculated that the effects of El Niño also influenced these results (Nur & Sydeman, 2009).
Aleesha Mattingly is a current student at The Evergreen State College, with a graduation date in June, 2019. Her studies mainly focus on wildlife biology, genetics, conservation and habitat preservation. She resides in Olympia, Washington.
Leave a Reply