

Order: Anseriformes
Family: Anatidae
Genus: Melanitta
Species: Melanitta perspicillata
Introduction
 Photo by Micah Riegner
Photo by Micah Riegner
 Photo by Micah Riegner
Photo by Micah Riegner
 Watercolor by Micah Riegner
Watercolor by Micah Riegner
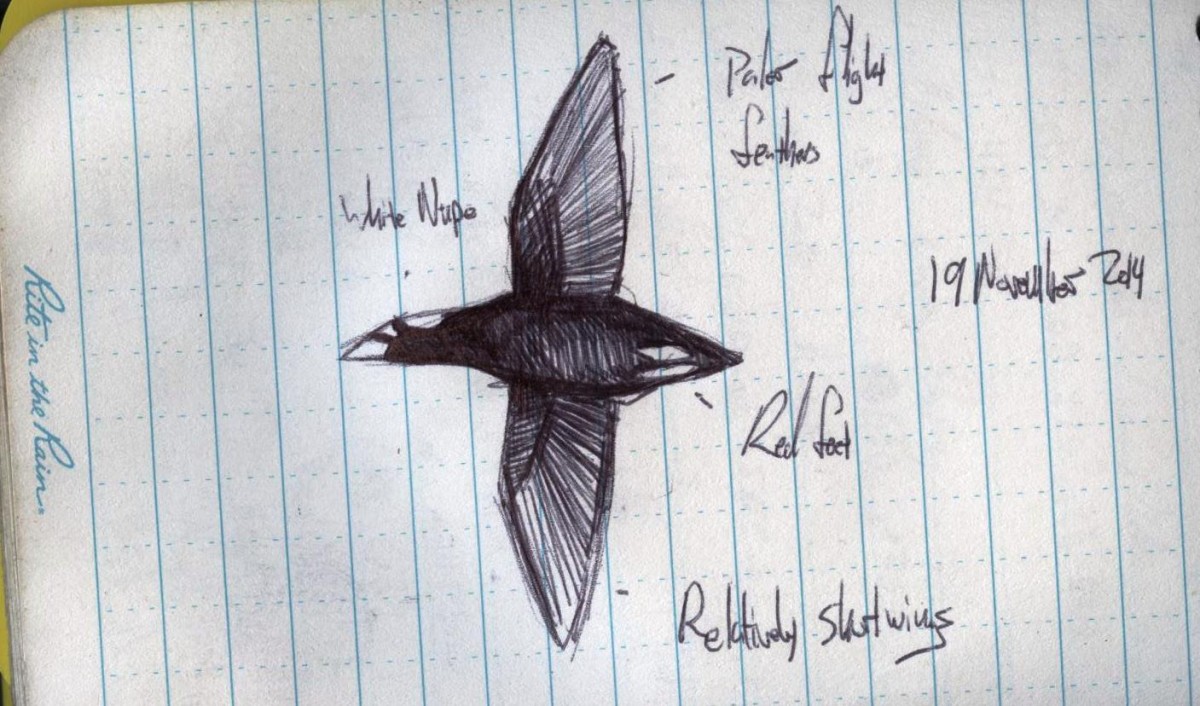 Field Sketch by Micah Riegner
Field Sketch by Micah Riegner
Surf Scoters are sea ducks endemic to North America. These molluscivores breed on lakes in the boreal forest, and winter on both Atlantic and Pacific Coasts (Savard et al. 1998, Takekawa 2011). Sea ducks as a group have been poorly studied, and Surf Scoters are one of the least-studied North American sea ducks (Wilson et al. 2010).
Males are black with distinct white patches on the forehead and back of nape: females are slightly smaller, and have dull gray plumage. They have relatively small wings for their body size, which results in high wing loading (Dickson et al. 2012). Both sexes exhibit the same bill morphology, but the males’ bills are bright orange and white (Savard et al. 1998). Immature males have limited white on the nape and the forehead (Sea Duck Information Series 2004). During the breeding season they eat freshwater invertebrates and in the winter, they feed primarily on mollusks, but will also take soft-bodied invertebrates (Anderson et al. 2008). They have been known to follow in the path of feeding Gray Whales to catch prey stirred up by the whales (Anderson et al. 2008). Western Surf Scoter populations have declined 60% in the past 30-50 years (Anderson et al. 2011). However, they are one of the few species that are benefiting from shellfish aquaculture (Faulkner 2012).
Surf Scoters breed along freshwater lakes in northern Canada and Alaska. Western, populations breed in inland Alaska, from the Kotzebue Sound to the Alaska Peninsula (Savard et al. 1998). Radio telemetry studies have shown that Surf Scoters have high nest site philopatry, and pairs arrive at the breeding sites by late May (Takekawa 2011). In Eastern Canada, the scoters breed around Quebec, however little research has been done in this area. (Savard et al.1998).
Surf Scoters winter along the Atlantic and Pacific coasts of North America. They winter from the Aleutian Islands to Sonora, Mexico on the Pacific Coast, and from Newfoundland to southern Florida, on the Atlantic Coast (Savard et al. 1998). The Alexander Archipelago has the highest densities of wintering Surf Scoters, but they are also abundant along the Coast of British Columbia (Savard et al. 1998). Studies using ratio telemetry have shown that Surf Scoters have high winter site fidelity and migration routes are determined by breeding sites (De La Cruz et al. 2012). They are the only species of scoter endemic to North America; the Black Scoter, and White-winged Scoters are also found in Eurasia. (Savard et al. 1998).
Climate change is expected to impact scoters especially on their breeding grounds in the northern boreal forest where temperatures are rising faster than in other areas (Takekawa 2011).
During the breeding season, Surf Scoters inhabit shallow fresh-water lakes in the northern boreal forest and tundra of northern Canada and Alaska (Savard et al. 1998, Takekawa 2011). Much of their breeding range is remote and difficult to survey (Sea Duck Information Series 2004). Recent studies have shown that Scoter densities on the breeding grounds are 0.11 pairs per km2 (Brook et al. 2012).
In the winter, Surf Scoters prefer shallow coastal waters, within 1 km of the shore (Savard et al. 1998). They will remain close to shore by day and move farther out by night (Lewis, et al. 2005). They will feed from mud, sand, and pebble substrates and are the only scoter that will inhabit rocky coasts (Savard, et al. 1998). They will switch from mussel beds in winter to eelgrass beds to feed on herring spawn in the early spring. Surf Scoters have more wintering habitat requirements than White-winged Scoters (Anderson et al. 2011). In the Puget Sound, Surf Scoters have a positive association with shellfish aquaculture (Faulkner 2012). .
During the winter, the Surf Scoter will feed predominantly upon mollusks, which they tear off the ocean substrate, but they will also consume crustaceans and soft-bodied prey items such as polychaetes (Anderson et al. 2008). They likely key in on the siphon holes to locate prey, but this behavior has yet to be observed (Lewis et al. 2005). As the winter progresses, they feed on fewer and fewer bivalves and their movements reflect prey availability (Anderson et al. 2009, Kirk 2007, Lewis et al. 2007). In areas with high prey availability, scoters spend less time foraging. Adults have been known to spend more time underwater than hatch year birds (Lewis et al. 2008).
Studies of stomach contents indicate that Surf Scoters have a diverse diet and they feed on smaller prey items than do White-winged Scoters (Anderson et al. 2008). They also have more microhabitat requirements and are less resistant to seasonal change than White-winged Scoters (Anderson et al. 2008, Anderson et al. 2011). In areas where scoters and scaup exist in the same habitat, scoters tend to take prey items that are buried deeper in the substrate (Richman and Lovvorn 2009).
Surf Scoters will modify seasonal movements in response to rich food resources (Lacrouix et al. 2005). During the spring migration, Surf Scoters will eat Pacific Herring eggs, and follow the spawning fish to take advantage of the rich food source (Lok et al. 2012). The herring eggs remain for 2 weeks before they hatch (Lacroix et al. 2005). Radio telemetry studies in Bayne Bay, British Columbia have shown that both Surf and White-winged Scoters time their northward migration to coincide with the Herring migration (Lok et al. 2008). Just as dabbling ducks follow the green wave of blooming algae, Scoters follow the northward wave of spawning herring (Lok et al. 2012). Scoters spend 50% less time underwater when feeding on herring eggs than when feeding on other prey. This provides them more time for courtship (Lewis et al. 2007). In 2003, a group of 5,000 Surf Scoters congregated off the coast of British Columbia to feed on Ophryotrocha polycheate. This remarkable congregation demonstrates how scoters will capitalize on an ephemeral resource (Lacrouix et al. 2005).
In the breeding season, adults feed a variety of aquatic insects and spiders to the young (Savard et al. 1998).
The Scoters will forage close to shore by day, and move out to sea by night as a predator-avoidance strategy. They have also been known to feed at night, though not as frequently as other sea ducks (Lewis et al. 2005). In the Straights of Georgia, British Columbia, Surf Scoters feed primarily on the Mussel Mytilus trossulus, which forms extensive beds along the rocky coast (Lacroix 1996). The scoters can strip a mussel bed in a very short period of time, due to a positive feedback loop; the more mussels are extracted from the beds, the easier it is to extract more mussels (Lacroix 1996). The introduced Varnish Clam Nuttallia obscura has increased food availability for Surf Scoters in British Columbia (Lewis et al. 2005). Clam densities plummet significantly during the winter when Surf Scoters are present. By the end of winter, much of the scoters’ food supply is depleted, and the birds’ body mass may drop over 9% (Lewis et al. 2007).
Wing Noise and Voice. Recorded on the Evergreen Beach on 16 November. The birds were about 20 meters away.
Wing Noise and Voice. Recorded on the Evergreen Beach on 5 December.
Scoter sounds underwater (also known as nothing). Recorded on 24 november.
Male Surf Scoters will display for females on their wintering grounds, and pairs will form before migration (Sea Duck Information Series 2004). The pairs will migrate to the breeding grounds where the males defend a moving territory: the females are not territorial (Savard et al. 1998).
Few nests have ever been described because of the remoteness of the breeding grounds. The few that have been described have been about 19 meters away from lake shores in the boreal forest. They were lined with down feathers and vegetation (Savard et al. 1998). Surf scoters have the shortest pair bond of any duck (Takekawa 2011). The female will spend 29 days incubating her 7-9 eggs. The male will leave the female once she starts incubating (Sea Duck Information Series 2007).
After the breeding season, the scoters will undergo a molt migration to remote coastal areas away from the breeding sites (Sea Duck Information Series 2004). They will molt the flight feathers before undergoing body molt and eclipse plumage (Savard et al. 2007). Male and females will migrate to different molting sites and the different age classes have a wide range of emergence dates. The remigial molt (molt of flight feathers) typically takes place between late June and September (Dickson et al. 2012). Researchers are working to better understand molt chronology in order to promote more efficient management practices (Savard et al. 2007).
Groups of Surf Scoters exhibit synchronous diving when feeding (Lewis et al. 2008, Schenkeveld et al. 1985). The reasons for synchronous diving are not entirely clear, though there is speculation that this behavior might increase foraging efficiency (Savard et al. 1998). It could also be an adaptation to avoid kleptoparasitism by gulls (Schenkeveld et al. 1985). Scoters have been known to follow Gray Whales to pick off the benthic organisms dislodged by the feeding whales. When the whales do suction sieving, they create large pits on the sea floor, which dislodges prey for sea ducks (Anderson et al. 2008).
Surf Scoters are unusual among ducks in being long lived with a low reproductive output. It takes them 2-3 years to reach sexual maturity (Sea Duck Information Series 2004). Surf Scoter surveys are difficult due to the remoteness of their breeding grounds, but the latest estimates suggest there are between 600,000- 1 million birds (Sea Duck Information Series 2004). According to the IUCN Red List of Endangered Species, the Surf Scoter is of least concern, but there has been a steady decrease in the western population in the last few decades. (BirdLife International 2012).
Out of the 15 species of sea duck, 10 are experiencing population declines, and it is estimated that Surf Scoters have declined 50 % in the west since the 1950s (Sea Duck Information Series 2004, Wilson et al. 2010). In the Puget Sound, both Surf Scoters and White-winged Scoters have declined by 60% in the past 30 -50 years (Anderson et al. 2011). A recent study suggests that pollutants in the water could be linked to Scoter mortality on their wintering grounds. Specimens collected off the coast of British Columbia had high levels of cadmium concentrations, which negatively influences vitamin A. These high levels of cadmium are bio accumulated through the food chain and could have negative health implications on breeding scoters (Wilson et al. 2010). Declines could also be linked to the fact that sport hunters shoot between 25,000 and 30,000 Surf Scoters per year (Sea Duck Information Series 2004).
Conservation efforts need to address multiple coastal habitats because Surf Scoters have diverse foraging preferences during the winter (Anderson et al. 2011,De La Cruz 2012). Climate change is expected to negatively influence Surf Scoters especially during the breeding season because the northern boreal forest is warming faster than other areas (Takekawa et al. 2011). While shellfish aquaculture has been known to negatively influence waterbirds, sustainable aquaculture has little effect on Surf Scoter populations in British Columbia and the Puget Sound (Ramūnas et al. 2006, Faulkner 2012). In some cases shellfish aquaculture has improved feeding habitat for wintering Surf Scoters because the mussels on aquaculture structures are larger and have a lower shell mass than wild ones (Kirk 2007).
Field Notes
8 November 2014
Evergreen Beach
47°04’52.3″N 122°58’56.4″W
Weather: 10% Cloud Cover; 0 Wind; 60 F
08.00 This was my first morning spent gathering foraging behaviors on the Scoters. There were 2 males and 1 female diving about 20 meters off shore. They were catching clams about 1 x bill length.
12 November 2014
Evergreen Beach
47°04’52.3″N 122°58’56.4″W
Weather: 10% Cloud Cover; 0 Wind; 60 F
14.00 There was a large group of Scoters diving about 10 meters off shore. I got some good dive times from them.
15.58 There were 5 male scoters fighting and doing the head bobbing display.
16 November 2014
Evergreen Beach
47°04’52.3″N 122°58’56.4″W
Weather: 10% Cloud Cover; 0 Wind; 65 F
14.25 I made some feeding observations of a group of scoters diving close to shore. They were diving around the submerged trees.
16.50 There were 2 male scoters displaying to a female. They were chasing each other making 10 meter flights and landing with their wings outstretched. They did the head bobbing displays, and made low clicking sounds, which were recorded.
17 November 2014
Evergreen Beach
47°04’52.3″N 122°58’56.4″W
Weather: 0% cloud cover, 0 wind, 65 F.
14.14 There was a single male scoter diving out in the middle of Eld inlet.
15.58 There was a huge raft of scoters displaying out by the dock on the north end of the Evergreen Beach. I counted 49 males chasing each other around and making low clicking sounds. Several of them would chase a single female, and she would often escape by diving.
19 November 2014
Evergreen Beach
47°04’52.3″N 122°58’56.4″W
Weather: 100% cloud cover, 0 wind, 50 F
15.00 I went down to record the diving scoters with Alison’s underwater microphone… much more easily said than done. The scoters were doing the cooperative diving out by the dock on the north end of the beach, but they were too far to record. My plan is to set the microphone hanging from the dock and let it record for an hour while the scoters feed.
15.11 The scoters stopped foraging, and gathered to display in the middle of the inlet. The males continued to chase the females. Out of the 77 males that were visible above water, 6 were in the midst of fighting or displaying.
15.18 Scoters mating! The male sat on top of the female to the point where she was almost completely underwater. This lasted for no more than 5 sec.
14.30 I watched 2 male scoters diving close to shore among the fallen tree trunks. There were several Barrow’s Goldeneyes diving with them.
20 November 2014
Nisqually National Wildlife Refuge
47°05’29.9″N 122°43’38.8″W
Weather: 100% cloud cover, 0 wind, 50 F
12.29 I watched a female scoter diving in the middle of the main channel and collected dive times. There was a Common Loon diving close by.
22 November 2014
Evergreen Beach
47°04’52.3″N 122°58’56.4″W
15.00 Kyle and I went down to the dock on the north end of the campus to try to record the scoters underwater. The scoters were diving close to the microphone, but there was too much static noise going on to hear anything else.
23 November 2014
Evergreen Beach
47°04’52.3″N 122°58’56.4″W
Weather: 100% cloud cover, 0 wind, 50 F, rain.
10.14 I spent several hours watching the scoters in the rain. I was watching a female (scoter) foraging. She was foraging alone and then a male joined her.
24 November 2014
Evergreen Beach
47°04’52.3″N 122°58’56.4″W
Weather: 100% cloud cover, 0 wind 60 F, rain.
10.00 I spent several hours down at the water trying to record the scoters. There was a small group diving around some fallen logs. When they were all underwater I approached them and submerged the microphone. I recorded some interspersed clicks that may or may not be the scoters.
2 December, 2014
Evergreen Beach
47°05’29.9″N 122°43’38.8″W
Weather: 0 % cloud cover, 0mph wind, 30 F
13.48 I spent the afternoon watching the scoters. There were 4 males and 4 females, which seemed to be paired up already. Two harbor seals came in and tried to catch the scoters, which promptly flew off.
14.40 There was a huge raft of scoters out at the north end of the Evergreen Beach, but they were too far to gather behavioral observations.
15.00 A large group was feeding very close to shore. I managed to get quite close to them by approaching them when they were underwater.
7 December, 2014
Evergreen Beach
47°05’29.9″N 122°43’38.8″W
Weather: 50% cloud cover, 0 mph wind, 35 F.
16.00 I watched a group of scoters that were close to shore. First there was a pair that was joined by another male. The three birds dived together and then one of the males flew off. Soon after that, a large raft of over 20 individuals came by and started feeding around the submerged trunks. After diving, they all began to preen in a line. The first bird would initiate the dive preening and the other birds would follow. I heard gunshots from the other side of the inlet, and the scoters departed.
16.29 Barrow’s Goldeneyes flew by. The wing noise was higher pitched than that of the Surf Scoters.
Surf Scoter Diving Behavior
Introduction
Synchronized diving has been observed in Surf Scoters and several groups of diving birds (Schenkeveld et al, 1985). During the winter, large groups of scoters will gather and dive together to capture mollusks from the sea floor. This behavior in which groups dive and surface at once has not been well studied (Savard et al. 1998). Some speculate that the scoters dive and surface together to avoid kleptoparisitism by gulls, or to find concentrated patches of mollusks on the sea floor (Schenkeveld et al 1985). The birds may also work together to dig up prey (Lewis et al. 2005). My question is whether scoters have a higher success rate in capturing prey items when they dive in groups as opposed to diving alone. I test this question using the null hypothesis there is no difference in success rates based on the number of diving scoters.
Methods
The Eld Inlet is narrow waterway located at the southern end of the Puget Sound 47°04’52.3″N 122°58’56.4″W. I did the majority of the field observations along the Evergreen Beach, where the Eld Inlet is about 1 mile wide and 46 feet deep. The substrate ranges from cobbles to coarse sand, and the water is generally calm. There are several submerged trunks along the coastline, and at high tide, the water can reach up to the forest.
I observed the feeding behavior of Surf Scoters between 8 November, and 2 December 2014. I walked along the Evergreen Beach to find diving scoters, estimated their distance from the shoreline, recorded the time they spent underwater, and how often they surfaced with food items. When I encountered scoters diving I watched them for as long as possible until they flew off, or started preening. When groups of Scoters did synchronous diving, I recorded the number of males and females that went down, and how many came up with food items. I recorded food items if they were identifiable however, it was often difficult to tell what the birds were eating if the food items were small.
Another part of the project was to get audio recordings of the scoters underwater using an underwater microphone. I made several attempts to record the scoters when they were diving close to shore and found that the best way to get close to them was to approach them while they were underwater. At times I was able to get within 5 meters of the birds.
The data were entered into excel, and statistical analyses were done on JMP pro. I ran a linear regression to see whether the group size and success rate fit a linear model. The other regressions I did compared dive time to distance from shore, number of attempts to group size, and group size to distance from shore.
Results
Surf Scoters were fairly common and easy to see during the time of the study. Most of the group diving attempts occurred close to shore while the single individuals were farther out (Fig 4). I recorded 155 diving events of individuals and groups ranging from 2 to 9 individuals.
Out of the 28 food items I was able to identify, 26 were clams, 1 was a crab, and 1 was a soft invertebrate (most likely a polycheate). The bivalves they were likely feeding on include Clinocardium nuttallii, Macoma sp., and Panopea generosa (Dethier 2006).
The linear regression showed that the success per attempt (success rate) increased significantly with a larger group size (Fig. 1). This was not related to increased attempts, which actually declined with group size. Success rate per individual also increased with larger group size, however, a linear model did not fit the data well. The data for this regression look to be normally distributed with the highest success occurring in groups of 4 individuals.
The recordings made with the underwater microphone did not confirm that scoters vocalize underwater.
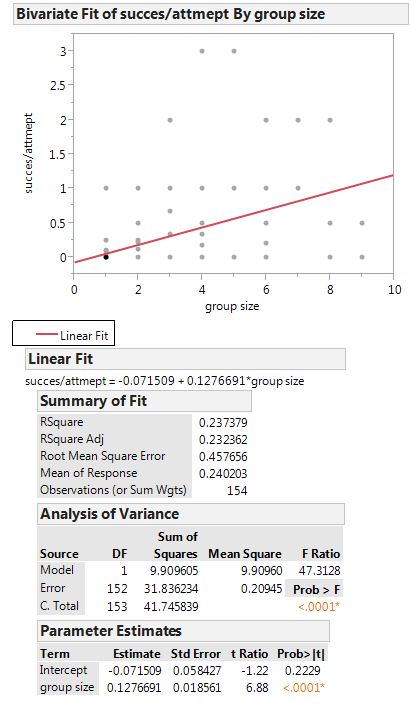
Figure 1. Linear regression showing increased success per attempt with increased group size. This came out to be significant with a P value of <0.0001. The r square value was 0.237379. I also looked at the relationship of increased group size and dive attempts and found that there was a decrease in attempts with larger group size. This means the increase in success rate is not related to increased attempts.

Figure 2. Map of the Evergreen Coast from google earth.
Discussion
Synchronized diving in Surf Scoters could be an adaptation to increase foraging efficiency, or to avoid kleptoparasitism by gulls (Schenkeveld et al. 1985, Lewis et al.). This study suggests that scoters increase their success rate when diving in groups. I speculate that they could be working together underwater to dig up benthic prey, or they could be alerting each other to rich food zones. Large groups feeding close to shore could also be a predator avoidance strategy of safety in numbers. The fact that Scoters are diving for longer periods in deeper water makes pretty logical sense.
Due to the fact that feeding behavior was relatively rare, whenever I encountered diving scoters I watched them for as long as possible. This caused my sample size for foraging observations of single individuals to be larger than that of large groups, which skewed the data. There would have been clearer results had I spent more time watching the large groups. I suggest that a future study invest more effort in making sure that sample sizes for each group are the same. Also, I suggest further research into underwater sound recording.
Anderson, E., J. R. Lovvorn, and M.T.Wilson. 2008. Reevaluating Marine Diets of Surf and White-Winged Scoters: interspecific differences and importance of Soft-bodied Prey The Condor110: 285-295.
Anderson, E. M., and J. R. Lovvorn. 2008. Gray whales may increase feeding opportunities foe avian benthivores. Marine Ecology Progress Series 360:291-296.
Anderson, E. M. 2009. Contrasts in nutrient metabolism and foraging strategies of Surf and White-winged scoters in near shore marine habitats. Ph.D. dissertation, University of Wyoming, Laramie.
Anderson, E. M., and J. R. Lovvorn 2011. Conrtasts in energy status and marine foraging strategies of White-winged Scoters (Melanita fusca) and Surf Scoters (Melanita perspicillata). The Auk. 128:248-257.
BirdLife International 2012. Melanitta perspicillata.. IUCN Red List of Threatened Species.
Brook, R., K. F. Abram, K. R. Middle, and R. K. Ross. 2012. Abundance and habitat selection of breeding Scoters (Melanitta spp.) in Ontario’s Hudson Bay Lowlands. The Canadian Field-Naturalist. 126: 20-27.
De la Cruz, S. E., J. Y. Takekawa, M. T. Wilson, D. R. Nysewander, J. R. Evenson, D. Esler, W. S. Boyd and D. H. Ward. 2009. Spring migration routes and chronology of surf scoters (Melanitta perspicillata): a synthesis of Pacific coast studies. Canadian Journal of Zoology. 87:1069-1086
Dethier, M. 2006. Native Shellfish in near shore Ecosystem of Puget Sound. Puget Sound Nearshore Partnership.
Dickson, R. D., D. D. Ester , J. W. Hupp , E. M. Anderson, J. R. Evenson, and J.J. Barrett. 2012. Phenology and duration of remigial moult in Surf Scoters (Melanitta perspicillata) and White-winged Scoters (Melanitta fusca) on the Pacific coast of North America. Canadian Journal of Zoology 90: 932-944.
Faulkner, H. 2013. Influence of Aquaculture on Winter Sea Duck Distribution and Abundance in South Puget Sound. Master’s Thesis. The Evergreen State College, Olympia.
Harris, M., L. Wilson, S. Trudeau, and J. Elliott. 2007. Vitamin A and Contaminant Concentrations in Surf Scoters (Melanitta Perspicillata) Wintering on the Pacific Coast of British Columbia, Canada. Science of The Total Environment 378 : 366-75.
Kirk, M. 2007. Movement and foraging behaviour of Surf Scoters wintering in habitats modified by shellfish aquaculture. M, Sc. Thesis. Simon Fraser University
Kirk M., D. Esler, and W. S. Boyd. 2007. Morphology and density of mussels on natural and aquaculture structure habitats: Implications for sea duck predators. Marine Ecology Progress Series 346:179-187.
Lacroix D. 1996. Foraging Impacts and Patterns of Wintering Surf Scoters Feeding on Bay Mussels In Coastal Strait of Georgia, British Columbia. Thesis Submitted for Master of Science at Simon Fraser University.
Lacroix, D. L., S. Boyd, D. Esler, M. Kirk, T. Lewis, and S. Liposvsky. 2005. Surf Scoters (Melanitta perspicillata) aggregate in association with ephemerally abundant polychaetes. Marine Ornithology 33:61-63.
Lewis, T. L., D. Esler, and W. S. Boyd. 2007. Effects of predation by sea ducks on clam abundance in soft-bottom intertidal habitats. Marine Ecology Progress Series 329:131–144.
Lewis, T. L., D. Esler, and W. S. Boyd. 2007. Foraging behaviors of Surf Scoters and White- winged Scoters during spawning of Pacific Herring. The Condor109: 216-222.
Lewis, T. L., D. Esler, W. Boyd. 2008. Foraging behavior of Surf Scoters (Melanitta perspicillata) and white-winged scoters (M. fusca) in relation to clam density: inferring food availability and habitat quality. Auk 125:149–157
Lewis, T. L, D. Esler, W. S. Boyd, and R. Zydelis. 2005. Nocturnal foraging behavior of wintering Surf Scoters and White-winged Scoters. Condor 107: 637-647
Lok, E.K., M. Kirk., D. Esler, and S.W. Boyd. 2008. Movements of Pre-Migratory Surf and White-Winged Scoters in Response to Pacific Herring Spawn. Waterbirds: The International Journal of Waterbird Biology 31: 385-393
Lok, E. K., D. Esler, , J. Y. Takekawa, De la Cruz, S. W., Boyd, W., Nysewander, D. R., & Ward, D. H. 2012. Spatiotemporal associations between Pacific herring spawn and surf scoter spring migration: evaluating a ‘silver wave’ hypothesis. Marine Ecology Progress Series 457:139-150.
Ramūnas, Ž. , D. Esler, W. S. Boyd, D. L. Lacroix and M. Kirk. 2006. Habitat use by wintering surf and white-winged Scoters: effects of environmental attributes and shellfish aquaculture. The Journal of Wildlife Management 70:1754-1762
Richman, S., and J. R. Lovvorn. 2009. Predator size, prey size and threshold food densities of diving ducks: does a common prey base support fewer large animals? Journal of Animal Ecology 78:1033-1042
Savard J.P., A. Reed, and L. Lesage. 2007. Chronology of breeding and molt migration in surf scoters (Melanitta perspicillata). Waterbirds: The International Journal of Waterbird Biology 30: 223-22
Savard J.P., D. Bordage and A. Reed. 1998. Surf Scoter (Melanitta perspicillata) The Birds of North America Online (A. Poole, Ed.). Ithaca: Cornell Lab of Ornithology; Retrieved from the Birds of North America Online:http://bna.birds.cornell.edu/bna/species/363
Sea Duck Information Series. 2004. Surf Scoter (Melanitta perspicillata). Sea Duck Joint Venture Online: http://sdjv.org
Schenkeveld, L., Ydenberg, R. 1985. Synchronous diving by surf scoter flocks. Canadian Journal of Zoology. 63(11):2516-2519
Takekawa, J., S. De La Cruz, M. T. Wilson, E. C. Palm, J. Yee, D. R. Nysewander, W. S. Boyd, and D. H. Ward. 2011. Breeding distribution and ecology of Pacific coast Surf Scoter. Boreal birds of North America: a hemispheric view of their conservation links and significance. Studies in Avian Biology. 41: 41-64
Vermeer, K. 1981. Food and populations of Surf Scoters in British Columbia. Wildfowl 32:107–116
Wilson, L. K., M. L. Harris, S. Tradeau , M. G.Ikonomou, , and J. E. Elliot. 2010. Properties of blood, porphyrins, and exposure to legacy and emerging persistent organic pollutants in Surf Scoters (Melanitta perspicillata) overwintering on the south coast of British Columbia, Canada. Archives of Environmental Contamination and Toxicology.59: 322–333
Leave a Reply