

Order: Strigiformes
Family: Strigidae
Genus: Glaucidium
Species: Glaucidium gnoma
Introduction and Description
Northern Pygmy-Owls are quirky little owls in my personal opinion, though the same can doubtless be said of other pygmy owls. They are cute, tiny, ferocious hunters. Unlike traditional owls they are diurnal (Giese & Forsman, 2003; Holt & Leroux, 1996; Jageman, Frye, & Garton, 2018; Sater, Forsman, Ramsey, & Glenn, 2006; Sibley, 2016) hunting by light rather than sound (Härmä, 2011; Sater et al., 2006). Patient birds they perch in daylight and wait, and wait, and wait, and pounce! They dive straight from their perch to their prey. (Jageman et al., 2018)
Northern Pygmy-Owls are small, quintessentially cute, owls that are mostly brown with white speckles or spots. The lower breast and belly are white with dark brown-black vertical streaks, the back of the head has two eyespots, and the tail is brown with white barring. The eyes are large and forward facing, the beak is sharply hooked, and the feet are raptorial. This description is based on photographs (Sibley, 2016) and a specimen from The Evergreen State College Natural History Museum (Specimen 3420) and photographs thereof (Wiklund, 2019a; Wiklund, 2019b).
- Length 6.75 in. (Sibley, 2016).
- Wing span 12 in. (Sibley, 2016).
- Mass ~67 g (average taken from the values provided by: Holt & Leroux, 1996; Jageman et al., 2018; Sibley, 2016).
There is a distinct lack of research on Northern Pygmy-Owls, as commented on by Jageman et al. (2018) and Giese and Forsman (2003) and as I personally observed, as such this page includes information on other pygmy owls as well in an attempt to extrapolate potential similarities among pygmy owls.

Northern Pygmy-Owl with purple mechanical pencil for scale. The specimen (3420) is from the Evergreen State College’s Natural History Museum and the photo is by Ælfhild Wiklund (2019b). Note the dark streaks on the breast and the raptorial feet and hooked beak. A vicious predator barely longer than my 6 in. pencil!

Northern Pygmy-Owl. The specimen (3420) is from the Evergreen State College’s Natural History Museum and the photo is by Ælfhild Wiklund (2019a). You can see the rough outline of the eyespots on the nape.
Northern Pygmy-Owls are found in western North America all the way from southern Alaska to Central America. They live in forested areas of the North American Pacific coast from Alaska to California and forested areas of the Rocky Mountains from British Columbia to Central America. (Jageman et al., 2018)
Northern Pygmy Owls are seasonally territorial and considered nonmigratory, though they might relocate to lower latitudes or lower elevations when confronted by cold weather and snow according to Hannah (1999) and Holt and Petersen (2000) as referenced by Jageman et al. (2018).
Northern Pygmy-Owls have been found to favor habitats possessing larger trees which are perforce older forests (Giese & Forsman, 2003; Jageman et al., 2018; Piorecky & Prescott, 2006; Sater et al., 2006) with greater structural diversity (Giese & Forsman, 2003; Piorecky & Prescott, 2006). One study (Jageman et al., 2018) found an association at one of their sites with riparian zones and another (Piorecky & Prescott, 2006) with wet moisture regimes in general. So, it seems old growth forests with running water would provide ideal habitat.
Trees prevalent in Northern Pygmy-Owl habitats include: Grand fir (Abies grandis), Douglas-Fir (Pseudotsuga menziesii), Western Red Cedar (Thuja plicata), White Pine (Pinus monticola), and Western Hemlock (Tsuga heterophylla) (Jageman et al., 2018); Lodgepole Pine (Pinus contorta), Douglas-Fir (Pseudotsuga menziesii), and Quaking Aspen (Populus temuloides) (Frye & Jageman, 2012); Bigleaf Maple (Acer macrophyllum), Red Alder (Alnus rubra), Douglas-Fir (Pseudotsuga menziesii), Western Red Cedar (Thuja plicata), and Western Hemlock (Tsuga heterophylla) (Deshler & Murphy, 2012); Douglas-Fir (Pseudotsuga menziesii) and Ponderosa Pine (Pinus ponderosa) (Sater et al., 2006); even aspen (Piorecky & Prescott, 2006). Sater et al. found Northern Pygmy-Owls to be habitat generalists in Oregon within conifer dominated or mixed conifer/hardwood forests (2006). Based on the dominant trees from the above studies it appears that conifers are the dominant vegetation in most Northern Pygmy-Owl habitats but that they are not overly particular birds and they also use mixed forest habitats or even, as per one location studied by Piorecky and Prescott (2006), deciduous dominated habitats in the case of the aspen.
Jageman et al. proposed that older forests with larger trees are beneficial in that they have more snags for nesting, canopies that provide cover with open understories to facilitate sit-and-wait hunting, and greater prey densities (2018). Piorecky and Prescott point out that these owls prefer larger habitat patch sizes (2006). This implies that Northern Pygmy-Owls may be more abundant in larger expanses of old growth forest.
The Evergreen State College campus forest is mostly mixed forest with a good deal of conifers and while it is not old growth it does have a good deal of decently aged tree-stands. Trees include: Western Red Cedar (Thuja plicata), Douglas-Fir (Pseudotsuga menziesii), Western Hemlock (Tsuga heterophylla), Bigleaf Maple (Acer macrophyllum), and Red Alder (Alnus rubra). This tree species assemblage aligns well with that found in studies of Northern Pygmy-Owls, as seen above. Furthermore, riparian zones are present on campus thanks to assorted small streams. This suggests that the Evergreen State College campus is a suitable habitat for Northern Pygmy-Owls.

The Evergreen State College campus forest. Forest stand with an open understory. Photo by Ælfhild Wiklund February 20th 2019.

The Evergreen State College Forest during the February 2019 snow event. Note the presence of both Western Red Cedar and Bigleaf Maple trees. Photo by Ælfhild Wiklund February 9th 2019.

The Evergreen State College campus forest during the February 2019 snow event. A creek undaunted by the storm. Photo by Ælfhild Wiklund February 9th 2019.
Northern Pygmy-Owls are sit-and-wait hunters; they remain stationary for extended periods of time and then pounce, performing rapid dives from their perch to their prey (Jageman et al., 2018; Holt & Leroux, 1996).
The Northern Pygmy-Owl’s prey consists of birds, mammals and insects (Frye & Jageman, 2012; Deshler & Murphy, 2012; Giese & Forsman, 2003). Additionally, I found one reference to the consumption of lizards reported by Holman 1926 (as cited by Holt & Leroux, 1996). These owls are considered food generalists according to Bent 1938, Holt and Leroux 1996, Giese 1999, Holt and Petersen 2000, and Giese and Forsman 2003 (as cited in Sater et al., 2006). I rather agree with that assessment as birds, mammals, and insects are very different animals, and then lizards too! It sounds like an extremely broad carnivorous diet.
Holt and Leroux said that they know of no other North American owl species with as high a percentage of birds in its diet as the Northern Pygmy-Owl (1996). However, overall these owls have a wide trophic niche and prey body mass ranged from 3 to 167g (average 38.4g) (Holt & Leroux, 1996). As the owl’s mass is about 67g (average taken from the values provided by: Holt & Leroux, 1996; Jageman et al., 2018; Sibley, 2016) that means that Northern Pygmy-Owls can take down prey more than twice their mass. All hail the cute and mighty hunting owl!
Northern Pygmy-Owl trill followed by song near Olympia. (Recording by Tayler Brooks, accessed via xeno-canto, license at https://creativecommons.org/licenses/by-nc-nd/2.5/)
Northern Pygmy Owl song near Olympia. (Recording by Andrew Spencer, accessed via xeno-canto, license at https://creativecommons.org/licenses/by-nc-nd/2.5/)
Northern Pygmy-Owls (Glaucidium gnoma) are also known as Mountain, Cape, or Guatemalan Pygmy-Owls and have been previously classified as Glaucidium hoskinsii, cobanense, or californicum (“Glaucidium,” n.d.) thus while these recordings were labeled as Glaucidium californicum (Spencer, n.d.; Brooks, n.d.) they would be more properly labeled today as Glaucidium gnoma.
Northern Pygmy-Owls have a variety of vocalizations. Adult Northern Pygmy-Owl sounds include their primary song which is a ‘toot song,’ a ‘trill call,’ a ‘chitter call,’ an alarm call, and a two-note call similar to their primary song but usually with a slightly higher-pitched second note which was used by males prior to prey deliveries in a study by Frye and Jageman along with the rest of the sounds (2012). On the other hand, while fledglings are highly vocal their vocalizations are limited to the ‘begging call’ (Frye & Jageman, 2012). Sibley describes the Northern Pygmy-Owl’s song as composed of monotonous repeated single or double toots and notes that the Pacific birds usually only sing single notes every 2 seconds or so which is slower than the birds of the Interior West and Mexico and the Northern Saw-whet (2016). This corresponds well with the above recordings, which were taken near Olympia in the Capitol State Forest, WA, USA (Spencer, n.d.; Brooks, n.d.).
Northern Pygmy-Owls are diurnal (Baron & Acorn, 1997; Deshler & Murphy, 2012; Giese & Forsman, 2003; Sibley, 2016), or diurnal/crepuscular to be specific (Frye & Jageman, 2012; Holt & Leroux, 1996; Sater et al., 2006). They have been observed to be sedentary at night (Jageman et al., 2018) even staying in the same tree until dawn (Frye & Jageman, 2012). According to one bird guide they are often seen perched on a snag during the day which is when they hunt (Baron & Acorn, 1997). This makes them an oddity among owls (though not unique) as owls are stereotypically nocturnal.
Northern Pygmy-Owls are visual hunters: they actually have symmetrical ears, they cannot achieve silent flight, and they hunt by day from high perches according to Holt and Petersen 2000 (as cited in Piorecky & Prescott, 2006). According to Norberg 1977 the diurnal Pygmy Owl (Glaucidium passerinum), which is a European species, also lacks the bilateral ear asymmetry that is so crucial for prey location by nocturnal owls (as cited in Härmä, Kareksela, Siitari & Suhonen, 2011). A study by Härmä et al. on these same owls (Glaucidium passerinum) found that they, like many diurnal raptors and unlike nocturnal owls, can detect ultraviolet light and use ultraviolet reflecting vole scent marks when hunting: the authors propose that other diurnal hunting owls can do likewise (2011). As a significant portion of the Northern Pygmy-Owl’s diet consists of voles (Holt & Leroux, 1996) I find it likely that it too utilizes ultraviolet light.
Norambuena and Muñoz-Pedreros propose that the Austral Pygmy Owl’s (Glaucidium nana) diurnal lifestyle, with peaks of activity mid-morning and evening, may be due to the high proportion of crepuscular/diurnal prey in the owl’s diet (2012). I think this correlation can be reasonably extended to Northern Pygmy-Owls as well given their diet, which features songbirds (Holt & Leroux, 1996; Sibley, 2016) among other things (Holt & Leroux, 1996), since songbirds tend to be diurnal.
Northern Pygmy-Owls are sit-and-wait hunters (Holt & Leroux 1996; Jageman et al., 2018) which I take to mean that they spend a significant portion of their waking hours perched and scanning for prey. Jageman et al. consider them to be human tolerant because of their lack of response to observers (2018) which implies to me that the mere presence of an observer is unlikely to startle them into flight. Between these two factors it seems likely that efforts to find Northern Pygmy-Owls visually will involve a lot of staring at small, cryptic, lumps on trees, a supposition supported by Baron and Acorn (1997). In these efforts it should be noted that the Northern Pygmy-Owl has a long tail (Baron & Acorn, 1997; Sibley, 2016) which it habitually flicks (Baron & Acorn, 1997) and often keeps slanted to the side when perched (Sibley, 2016). So, if you set off in search of this owl, good luck!

Northern Pygmy-Owl (Glaucidium gnoma) chicks. Photo by Gail Hampshire on the 5th June 2013. Accessed via Flickr. The license is at https://creativecommons.org/licenses/by/2.0/deed.en
Northern Pygmy-Owls are cavity nesters (Deshler & Murphy, 2012; Giese & Forsman, 2003; Holt & Leroux, 1996).
A study by Deshler and Murphy (2012) on Northern Pygmy-Owls found:
- Clutch sizes ranged from 4-7 eggs.
- Incubation ~ 30 days.
- Nestling period ~27 days.
Deshler and Murphy also found that nestling survival was high, nest failure low, and offspring survival in the weeks following fledging high, therefor they concluded that Northern Pygmy-Owls were successful breeders in their study (2012).
Fledging is synchronous in Northern Pygmy-Owls (Frye & Jageman, 2012; Giese & Forsman, 2003): one study places it between June 18th and August 5th (Frye & Jageman, 2012) and another between mid-June and mid-July (Giese & Forsman, 2003).
Frye and Jageman studied Northern Pygmy-Owl family groups with a focus on the post-fledging dependency period. They found that males rarely delivered prey directly to the young but rather to the female and the female passed it on along with the prey she caught. That said males attended the fledglings longer than females. 1-10 days after the adult male left the young initiated natal dispersal. After that both adult males and adult females returned to the nest stand 6-35 days after leaving their young and mated pairs were often observed together once the female (who stayed independent longer) returned. While the family groups were still together the researchers observed allopreening between siblings and between adults and young. In observation of fledglings hunting the authors note that while focusing on prey and perches young owls used exaggerated head motions sometimes flipping their heads almost upside down: nonetheless 47% of fledgling hunting attempts observed were successful. (2012)
Small Northern Pygmy-Owl females, rather than large, breed earliest and sometimes lay larger clutches (Deshler & Murphy, 2012). Deshler and Murphy suggest that this occurrence may be due to the fact that smaller individuals require less food for maintenance and therefor can put more energy into breeding (2012). This may be an evolutionary mechanism that helps keep Northern-Pygmy Owls (and other pygmy owls) small.
Cavity Nesters
Northern Pygmy-Owls are cavity nesters (Deshler & Murphy, 2012; Giese & Forsman, 2003; Holt & Leroux, 1996) who frequently nest in woodpecker cavities (Giese & Forsman, 2003; Holt & Leroux, 1996). While they do not usually reuse nest cavities they do usually nest in the same areas each year (Deshler & Murphy, 2012).
Beyond Glaucidium gnoma, pygmy owls in general primarily nest in woodpecker cavities according to Voous 1998 (as cited in Sater et al., 2006). A study in southern Finland on Three-toed Woodpecker cavity use found that 26-31% of breeding attempts by Pygmy Owls (Glaucidium passerinum) were in old Three-toed Woodpecker cavities (Pakkala, Tiainen, Piha & Kouki, 2018). The Pygmy Owl also uses nest cavities originally created by other woodpeckers such as Great Spotted and Grey-Headed Woodpeckers, however it uses Three-toed Woodpecker cavities more than random chance allows for given the lower availability of these cavities compared to those made by Great Spotted Woodpeckers (Pakkala et al., 2018). The Eurasian Pygmy Owl (Glaucidium passerinum by another name) was found in the southern French Prealps to use nest cavities created by Great Spotted Woodpeckers (Dendrocopos major) (Barbaro et al., 2016). This implies to me that the distribution and density of cavity excavating woodpeckers, and the species of those woodpeckers, are key considerations in the study of Northern Pygmy-Owls.
Northern Pygmy-Owls do not, of course, exist in isolation. They interact with other birds in a number of ways. Most obviously they kill and eat songbirds (Holt & Leroux, 1996; Sibley, 2016). Also, they are the smallest of the Pacific Northwest coastal owls (Baron & Acorn, 1997) and are themselves prey for bigger raptors (Jageman et al., 2018). A study in Europe on spatio-temporal patterns within the owl guild found that the pygmy owl (presumably Glaucidium passerinum but I am unsure) may avoid territories controlled by the Ural owl due to predation of Ural owls on pygmy owls, however the authors note that other factors may explain the results (Kajtock, Matysek & Figarski, 2016). Thus, pygmy owls in general have an eat-and-be-eaten relationship with certain other members of class Aves.
One of the most prominent features of the Northern Pygmy-Owl in my mind are the eyespots. Eyespots are a common feature of all pygmy owls (Sibley, 2016). Deppe et al. hypothesized that the Northern Pygmy-Owl’s eyespots serve both anti-predator and anti-mob purposes. Their study found that mob birds avoided passing behind models of Northern Pygmy-Owls with eyespots. The authors hypothesize that this may have a two-fold advantage for pygmy-owls in that it may reduce attack from behind on pygmy-owls and increase the opportunities to turn mob birds into prey birds because they pass in front of the pygmy-owls instead. (2003) Baron and Acorn also suggest that the eyespots help intimidate mob birds (small songbirds) (1997). This suggests that eyespots are an evolutionary trait that helps protect Northern Pygmy-Owls from mobbing among other things.
Population Trends
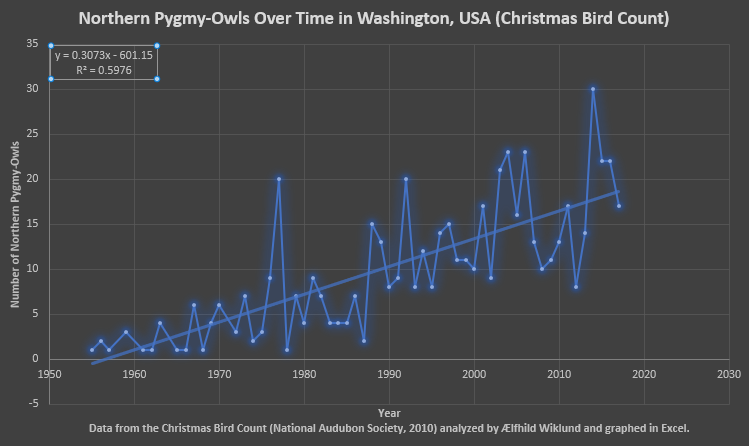
Fig. 1 Based on the data from the Christmas Bird Count for Washington State (National Audubon Society, 2010) over the last half century plus there has been an increase in the number of Northern Pygmy-Owls surveyed. The R2 value is ~0.6 which isn’t too bad especially compared to the graphs in the figures below. While this may represent survey method and observer bias it might correspond to an increase in their population.
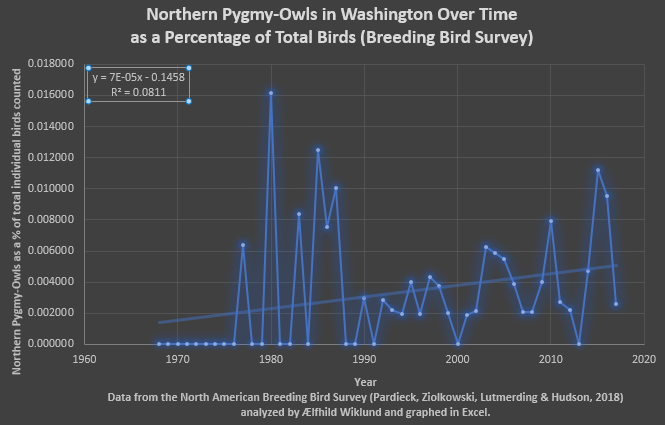
Fig. 2 Based on the data from the North American Breeding Bird Survey for Washington State (Pardieck, Ziolkowski, Lutmerding & Hudson, 2018) the percentage of individual birds counted that is composed of Northern Pygmy-Owls has gone up very slightly (though the R2 value, a mere ~0.08, suggests this is not a terribly significant correlation the trend is nonetheless present). I do not know if the initial sequence of zeros represents an absence of owls during the surveys or if the observers weren’t looking for them so I am uncertain if this represents observer bias. Since this is a percentage of the total birds surveyed it should account for bias in the form of observer effort to find birds in general and thus is more likely to actually represent an (extremely slight) increase in the Northern Pygmy-Owl population over the last half century.
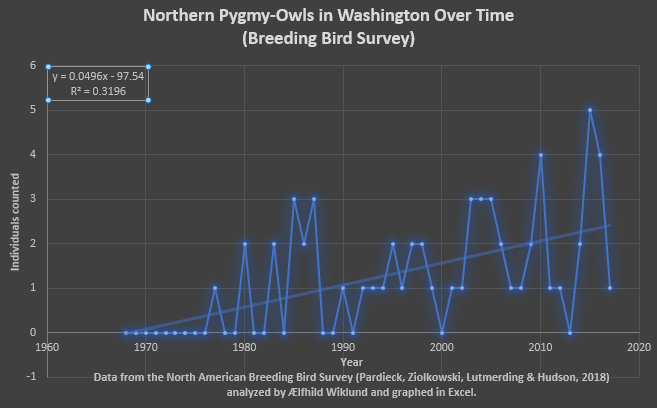
Fig. 3 Based on the data from the North American Breeding Bird Survey for Washington State (Pardieck, Ziolkowski, Lutmerding & Hudson, 2018) the absolute number of Northern Pygmy-Owls surveyed has gone up with an R2 value ~0.32 which is a stronger correlation than that in Fig. 2 a fact that may be due to increased observer effort in recent years, or not. I do not know if the initial sequence of zeros represents an absence of owls during the surveys or if the observers weren’t looking for them so I am uncertain if this represents observer bias. Given the lesser correlation in Fig. 2 I do not find this compelling evidence of a population increase of note.
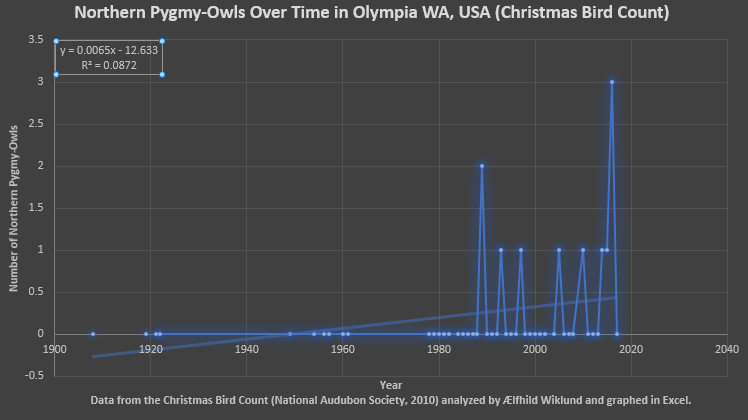
Fig. 4 Based on the data from the Christmas Bird Count for Olympia, WA (National Audubon Society, 2010) over the last century plus there has been an increase in Northern Pygmy-Owls counted. That said the data points for the first few decades are few and far between, the R2 is less than 0.09, and there have only been a few cases of even one owl being seen. Again, how much is observer bias I cannot say and the low quantities of owls observed make me suspect that this trend could be an artificial production of limited data mostly because of those zeros. Were the birds not seen because no one was looking for them, or because they weren’t in the right area, or because they weren’t there?
A few trends show when all four figures are considered together. In Figures 1-4 I observe that the quantity of Northern Pygmy-Owls surveyed undergoes fluctuations, often drastic, and is frequently zero. Whether this represents fluctuations in population or years when the surveyors simply didn’t observe this sparsely distributed and subtle bird I cannot say. Now I have enumerated all the reasons I think the apparent increase in population could be a mirage, however, I would like to point out that in all four sets of data analyzed there is an increase in Northern Pygmy-Owls over time, very slight and with poor correlation, but there: none of these graphs show a decrease when all the years are included. None of my research discussed population trends and based on the data from the North American Breeding Bird Survey (Pardieck, Ziolkowski, Lutmerding & Hudson, 2018) and the Christmas Bird Count (National Audubon Society, 2010) I cannot state anything conclusively but there may be a general increase in population over the last few decades. Furthermore, all four graphs from two data sets (and one subset, Olympia) show that the most recent trend (the last few years) is the decline following the latest peak in Northern Pygmy-Owls surveyed.
Conservation Issues
My research did not uncover any pressing conservation issues. That said I note that Northern Pygmy-Owls tend to be associated with larger trees and perforce older forests (Giese & Forsman, 2003; Jageman et al., 2018; Piorecky & Prescott, 2006; Sater et al., 2006) and greater structural diversity (Giese & Forsman, 2003; Piorecky & Prescott, 2006) all of which spells old growth forest from my perspective. Jageman et al. propose that older forests with larger trees are beneficial in that they have more snags for nesting, canopies that provide cover with open understories to facilitate hunting, and greater prey densities (2018). Piorecky and Prescott point out that Northern Pygmy-Owls prefer larger habitat patch sizes (2006) and Frye and Jageman found that during the post-fledging dependency period the family groups used areas ranging from 34.6-94.6 ha (2012) which means an even larger area or access to another area is needed for dispersal. Thus, I conclude that Northern Pygmy-Owl populations are likely to diminish with the loss of old growth forests even when the loss is merely diminishment in size. As the conservation of old growth forests in the Pacific Northwest of America is an ongoing issue it is clear that it should be kept in mind by those studying Northern Pygmy-Owls and vice versa.
A study by Barbaro et al. in France on Eurasian Pygmy Owls (Glaucidium passerinum) found that they preferentially select old-growth fir-dominated mixed forests and that the owl’s nesting cavities are associated with dead or decaying spruces. The authors conclude that these forests are key to the conservation of Eurasian Pygmy Owls at the lower latitudes of their range. (2016) The study was done at 44°56’N (Barbaro et al., 2016) which isn’t too much farther south than 47°N (Olympia) and so I consider this additional evidence that conservation of old growth forests in the Pacific Northwest of America may benefit Northern Pygmy-Owls. Barbaro et al.’s study also found that breeding adult male Eurasian Pygmy Owls (Glaucidium passerinum) have home ranges of 0.46-0.98 km2 with a mean of 0.67 km2 (2016). This is similar to the 34.6-94.6 ha areas used by family units of Northern Pygmy-Owls (Jageman & Frye, 2012) and provides further numerical support that large areas are needed.
Arndt, D. (2018). Northern Pygmy-Owl [Photo]. Retrieved from https://www.flickr.com/photos/ubermoogle/39248366504. License at https://creativecommons.org/licenses/by-nc-sa/2.0/legalcode
Barbaro, L., Blache, S., Trochard, G., Arlaud, C., Lacoste, N., & Kayser, Y. (2016). Hierarchical habitat selection by Eurasian Pygmy Owls Glaucidium passerinum in old-growth forests of the southern French Prealps. Journal of Ornithology, 157(1), 333–342. https://doi.org/10.1007/s10336-015-1285-3
Baron, N., Acorn, J. (1997). Birds of the Pacific Northwest Coast. Auburn, WA: Lone Pine Publishing.
Brooks, T. (n.d.). XC41302: Northern Pygmy Owl (Glaucidium californicum grinnelli) [Audio file]. Retrieved from https://www.xeno-canto.org/. License at https://creativecommons.org/licenses/by-nc-nd/2.5/
Deppe, C., Holt, D., Tewksbury, J., Broberg, L., Petersen, J., & Wood, K. (2003). Effect of Northern Pygmy-Owl (Glaucidium gnoma) eyespots on avian mobbing. The Auk, 120(3), 765–772. https://doi.org/10.1642/0004-8038(2003)120[0765:EONPGG]2.0.CO;2
Deshler, J. F., & Murphy, M. T. (2012). The breeding biology of the Northern Pygmy-Owl: Do the smallest of the small have an advantage? The Condor; Waco, 114(2), 314–322. http://dx.doi.org/10.1525/cond.2012.100140
Frye, G. G., & Jageman, H. R. (2012). Post-fledging ecology of Northern Pygmy-Owls in the Rocky Mountains. The Wilson Journal of Ornithology; Fort Collins, 124(2), 199–207.
Giese, A. R., & Forsman, E. D. (2003). Breeding season habitat use and ecology of male Northern Pygymy-owls. Journal of Raptor Research, 37(2), 117–124.
Glaucidium [gnoma, hoskinsii, cobanense or californicum] (Mountain, Cape, Guatemalan or Northern Pygmy-Owl) – Avibase. (n.d.). Retrieved March 13, 2019, from https://avibase.bsc-eoc.org/species.jsp?lang=EN&avibaseid=8D2303B04B4BE58C&sec=taxontable&version=aou&lang=EN&avibaseid=8D2303B04B4BE58C&sec=taxontable&version=aou
Härmä, O., Kareksela, S., Siitari, H., & Suhonen, J. (2011). Pygmy owl Glaucidium passerinum and the usage of ultraviolet cues of prey. Journal of Avian Biology, 42(1), 89–91. https://doi.org/10.1111/j.1600-048X.2010.05192.x
Hampshire, G. (2013). Northern Pygmy Owl chicks.Glaucidium gnoma [Photo]. Retrieved from https://commons.wikimedia.org/wiki/File:Northern_Pygmy_Owl_Chicks.Glaucidium_gnoma_-_Flickr_-_gailhampshire.jpg. License at https://creativecommons.org/licenses/by-nc-sa/2.0/legalcode
{kind=link}
Holt, D. W., & Leroux, L. A. (1996). Diets of Northern Pygmy-Owls and Northern Saw-Whet Owls in West-Central Montana. The Wilson Bulletin, 108(1), 123–128.
Jageman, H. R., Frye, G. G., & Garton, E. O. (2018). Habitat selection by Northern Pygmy-Owls in the Rocky Mountains. Journal of Raptor Research, 52(3), 349–361. https://doi.org/10.3356/JRR-17-21.1
Kajtoch, Ł., Matysek, M., & Figarski, T. (2016). Spatio-temporal patterns of owl territories in fragmented forests are affected by a top predator (Ural owl). Annales Zoologici Fennici, 53(3–4), 165–174. https://doi.org/10.5735/086.053.0405
National Audubon Society (2010). The Christmas bird count historical results [Online]. Available from http://www.christmasbirdcount.org [March-12-2019]
Norambuena, H. V., & Muñoz-Pedreros, A. (2012). Diurnal activity of the Austral Pygmy Owl (Glaucidium nana) in Southern Chile. The Wilson Journal of Ornithology; Fort Collins, 124(3), 633–635.
Pakkala, T., Tiainen, J., Piha, M., & Kouki, J. (2018). How important are nest cavities made by the Three-Toed Woodpecker Picoides tridactylus for cavity-nesting forest bird species? Acta Ornithologica, 53(1), 69–80. https://doi.org/10.3161/00016454AO2018.53.1.007
Pardieck, K.L., Ziolkowski Jr., D.J., Lutmerding, M., & Hudson, M.-A.R. (2018). North American breeding bird survey dataset 1966 – 2017, version 2017.0. U.S. Geological Survey, Patuxent Wildlife Research Center. https://doi.org/10.5066/F76972V8.
Piorecky, M. D., & Prescott, D. R. C. (2006). Multiple spatial scale logistic and autologistic habitat selection models for Northern Pygmy Owls, along the eastern slopes of Alberta’s Rocky Mountains. Biological Conservation, 129(3), 360–371. https://doi.org/10.1016/j.biocon.2005.11.003
Robledo-Campos, D. (2019). Ælfhild Wiklund at McLane Creek [Photo].
Sater, D. M., Forsman, E. D., Ramsey, F. L., & Glenn, E. M. (2006). Distribution and habitat associations of Northern Pygmy-Owls in Oregon. Journal of Raptor Research, 40(2), 89–98. https://doi.org/10.3356/0892-1016(2006)40[89:DAHAON]2.0.CO;2
Sibley, D.A. (2016). The Sibley field guide to birds of Western North America, (2nd ed.). New York, NY: Alfred A. Knopf.
Spencer, A. (n.d.). XC76354: Northern Pygmy Owl (Glaucidium californicum grinnelli) [Audio file]. Retrieved from https://www.xeno-canto.org/. License at https://creativecommons.org/licenses/by-nc-nd/2.5/
Wiklund, Æ. F. (2019a). Northern Pygmy-Owl: specimen (3420) is from the Evergreen State College’s Natural History Museum [Photo].
Wiklund, Æ. F. (2019b). Northern Pygmy-Owl with purple mechanical pencil for scale: specimen (3420) is from the Evergreen State College’s Natural History Museum [Photo].
Wiklund, Æ. F. (2019c). The Evergreen State College forest creek [Photo].
Wiklund, Æ. F. (2019d). The Evergreen State College forest stump in cedar stand [Photo].
Wiklund, Æ. F. (2019e). The Evergreen State College forest with open understory [Photo].
Acknowledgements
I would like to formally acknowledge the contributions of the vast number of people who contribute to online databases including the following. Thousands of U.S. and Canadian participants in the North American Breeding Bird Survey. Everyone who bundles up and heads out for the Audubon Society’s Christmas Bird Count. Members of the xeno-canto community including Andrew Spencer and Tayler Brooks. Members of the Flickr community including Daniel Arndt. Furthermore, I would like to thank the North American Breeding Bird Survey and the Audubon Society’s Christmas Bird Count for making their data readily available to the public as it was the foundation of my population analyses and graphs.
The author of this page, Ælfhild Wiklund, is a senior at the Evergreen State College pursuing a Bachelor of Arts and Science with a focus on environmental science. She is also a published poet. Ælf loves field science whether it involves peering at ducks from a cold beach or at a lichen on a nearby tree. She is a bit of a night owl and thus her choice to study one of the few diurnal owls is a bit ironic, to the great amusement of her mother. This project has made her even more intrigued by owls and brought new questions to mind. What makes an owl an owl? Is it appearance or behavior or genetics? The dichotomy of diurnal and nocturnal owls raises interesting evolutionary questions.

Photo of Ælfhild Wiklund taken by Dania Robledo-Campos at McLane Creek 20th February 2019.
July 6, 2021 at 10:00 pm
Hi,
The gap in information about northern pygmy-owls that you mention in your account has been closed somewhat by my research and I think it would help inform your account.
My manuscripts on pygmy-owls are the source for the best available information on the species’ clutch size, timing of breeding, magnitude of breeding, nest success (and failure), body size of the sexes, diet, prey composition, annual variation in diet, seasonal variation in diet, nest trees, nest site habitat, renesting following mate loss and nest loss, single-parenting, nest predators etc etc.
The longevity record for the species also comes from my dataset – two males that I captured and recaptured were at least four years old.
I am also the only researcher who has collected data on the northern pygmy-owl’s daily activity pattern, to demonstrate (with a lot of data on just one pair), rather than merely claim, that pygmy-owls are diurnally active, not crepuscular (as some have said), and how the timing of their activity relates to their diet during the breeding season.
Incidentally, pygmy-owls are not sit and wait predators, as you mention in your account. They are active predators that hunt by sight and sound, based solely on my many years of observation of them at and near their nests.
I hope to be revising the Cornell Bird’s of the World species account soon.
Good luck with your account. I hope my data can make it more accurate.
Cheers,
John Deshler
Please find:
Deshler JF. 2021. Northern Pygmy-Owl (Glaucidium gnoma) nesting ecology in northwestern Oregon. Wilson Journal of Ornithology 132(2):352-365.
https://doi.org/10.1676/1559-4491-132.2.352.
Deshler JF. 2021. Higher reversed sexual size dimorphism among nesting pairs of Northern Pygmy-Owls (Glaucidium gnoma) in northwestern Oregon than among specimens collected at the range-wide scale. Wilson Journal of Ornithology 132 (3): 619–627.
Deshler JF, Forsman ED, Swingle JK. 2019. Diurnal activity
pattern of a pair of breeding Northern Pygmy-Owls.
Northwestern Naturalist. 100:176–185.
Deshler JF, Murphy MT. 2012. The breeding biology of the
Northern Pygmy-Owl: Do the smallest of the small
have an advantage? Condor. 114:314–322.
Deshler JF. 2010. Nest-site habitat selection and breeding
biology of the Northern Pygmy-Owl in northwestern
Oregon [master’s thesis]. Portland (OR): Portland State
University.
March 24, 2022 at 8:08 pm
Thank you for all of this great information. We are going to remove that post for now. This blog is a student-driven project for students in my ornithology programs. Some of the species accounts are not as complete as others. I appreciate your response and feel free to contact me if you have other comments/questions (styringa@evergreen.edu). Alison Styring, Ph.D. Faculty